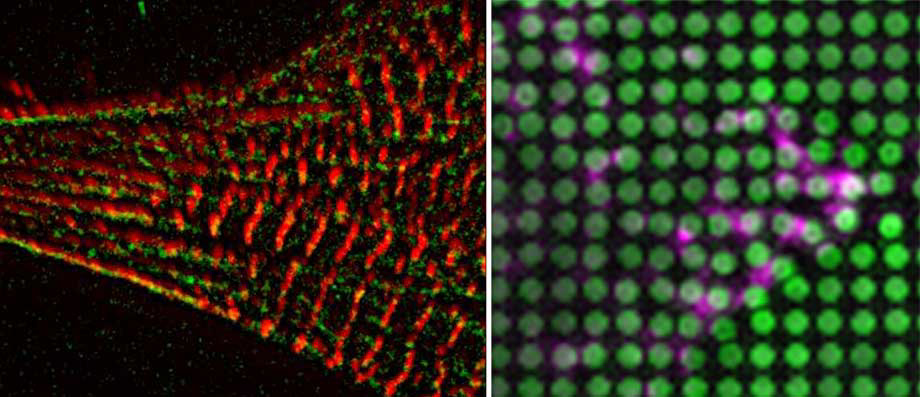
Cardiomyocyte Mechanosensing and Mechanical Imprinting in Heart Disease
Cells in the myocardium are exposed to stretching forces from hemodynamic pressure or the cardiomyocyte contraction as well as passive elasticity from the extracellular matrix (ECM), all of which are subject to change during development and disease.
Especially cardiac diseases, such as hypertrophic cardiomyopathies are frequently associated with myocardial fibrosis, which stiffens the mechanical environment of the cardiomyocytes.
While several studies have investigated the role of matrix stiffness on contractile properties of cardiomyocytes it is still unclear how this is influencing disease progression by altering mechanosignalling pathways in cardiomyopathies or after cardiac infarction.
Moreover the molecular mechanism behind cardiomyocyte rigidity sensing are still elusive. Here we aim to address these critical questions, using neonatal rat and hIPS cardiomyocytes, together with tools to apply or measure forces as well as live- (superresolution) microscopy.
Especially cardiac diseases, such as hypertrophic cardiomyopathies are frequently associated with myocardial fibrosis, which stiffens the mechanical environment of the cardiomyocytes.
While several studies have investigated the role of matrix stiffness on contractile properties of cardiomyocytes it is still unclear how this is influencing disease progression by altering mechanosignalling pathways in cardiomyopathies or after cardiac infarction.
Moreover the molecular mechanism behind cardiomyocyte rigidity sensing are still elusive. Here we aim to address these critical questions, using neonatal rat and hIPS cardiomyocytes, together with tools to apply or measure forces as well as live- (superresolution) microscopy.
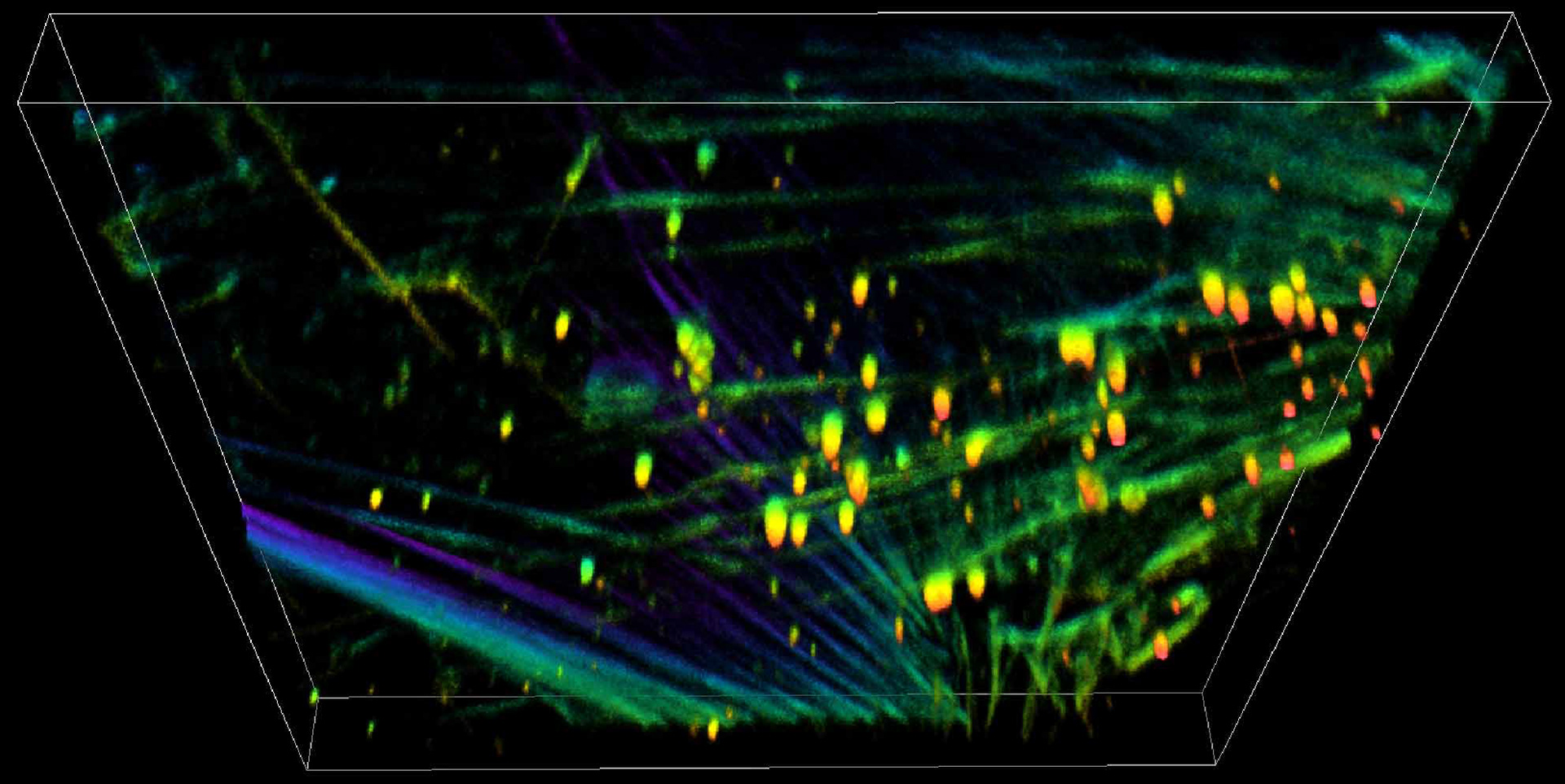
Vascular Smooth Muscle Cell Mechanosensing in Arterial Disease
Vascular smooth muscle cells (VSMC) take a central role in the onset and progression of many cardiovascular diseases, from atherosclerosis to vascular injury or restenosis, where their migration and matrix degradation function is deregulated. While generally responsible for maintaining the contractile tonus of the vessel walls and regulating the blood flow, VSMCs lose their contractility in response to the vascular injury, or during the disease progression and switch to a migratory phenotype. In addition to chemical signals from endothelial cells, VSMCs respond to mechanical stimuli. We are studying how different mechanical stimuli combine to drive the phenotypic change and arterial disease progression.
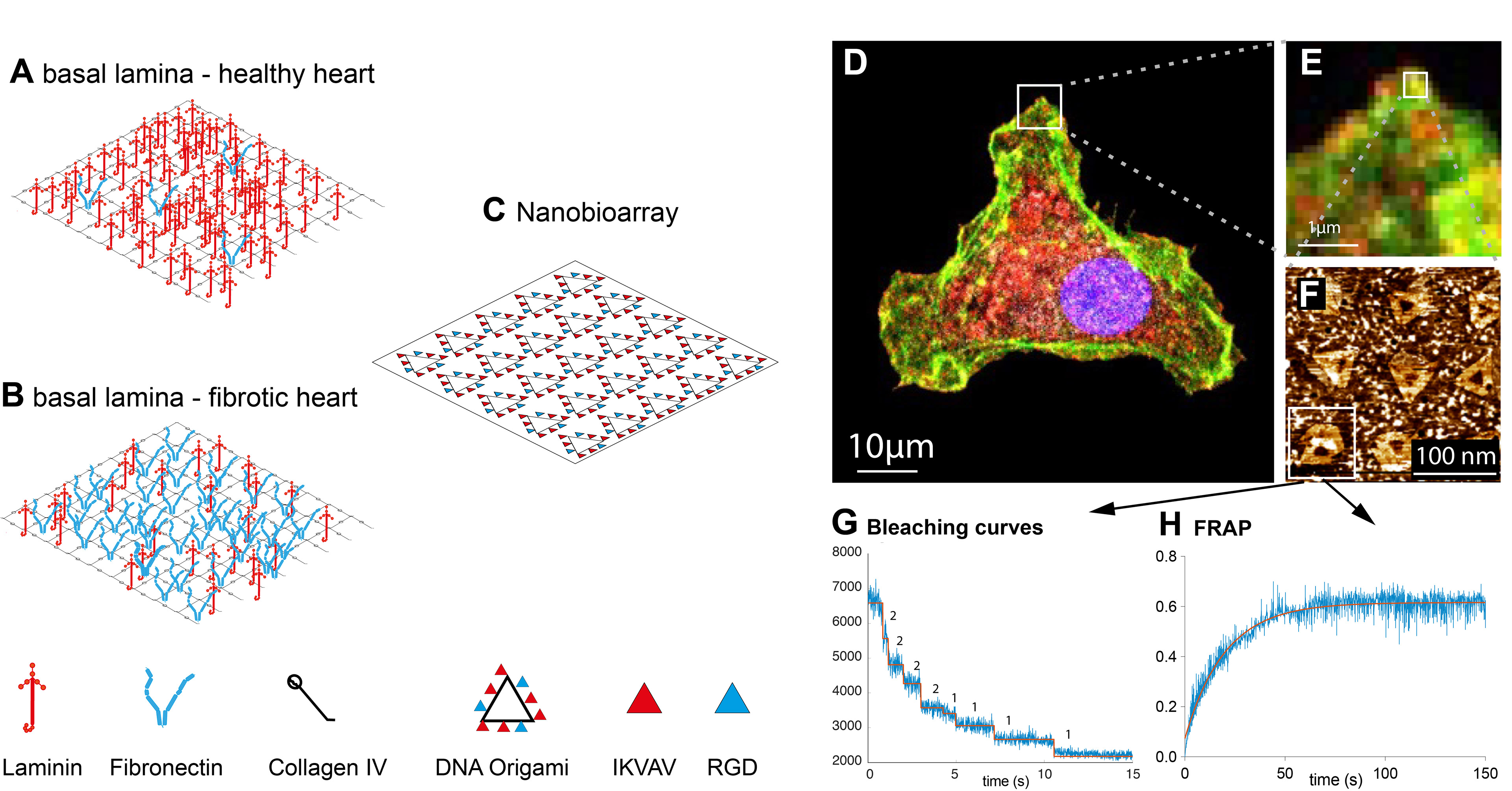
Nanopatterning for (Integrin) Adhesion Signalling
Integrins are heterodimeric transmembrane receptors that bind to the extracellular matrix. They are an integral part of focal adhesions, where they participate in inside-out and out-side in (mechano-)signalling. The composition of the integrin dimer determines the specificity towards either collagen, laminin or fibronectin peptide sequences. Amongst others, integrin mechanosignalling has been associated with the development of contractile structures in the heart during development, or pathological remodelling in heart disease. Previous studies indicated essential differences in the dynamics and mechanical properties of various integrins. However it is still unclear how the different cardiomyocyte integrins determine downstream signalling and force dynamics to influence cardiomyocyte differentiation, myofibrillar assembly and maintenance, or remodelling.
Together with our collaborators we aim to develop nanopatterning and DNA-origami techniques for defined ligand geometries and stochiometries for the analysis of the downstream signalling pathways.
Together with our collaborators we aim to develop nanopatterning and DNA-origami techniques for defined ligand geometries and stochiometries for the analysis of the downstream signalling pathways.
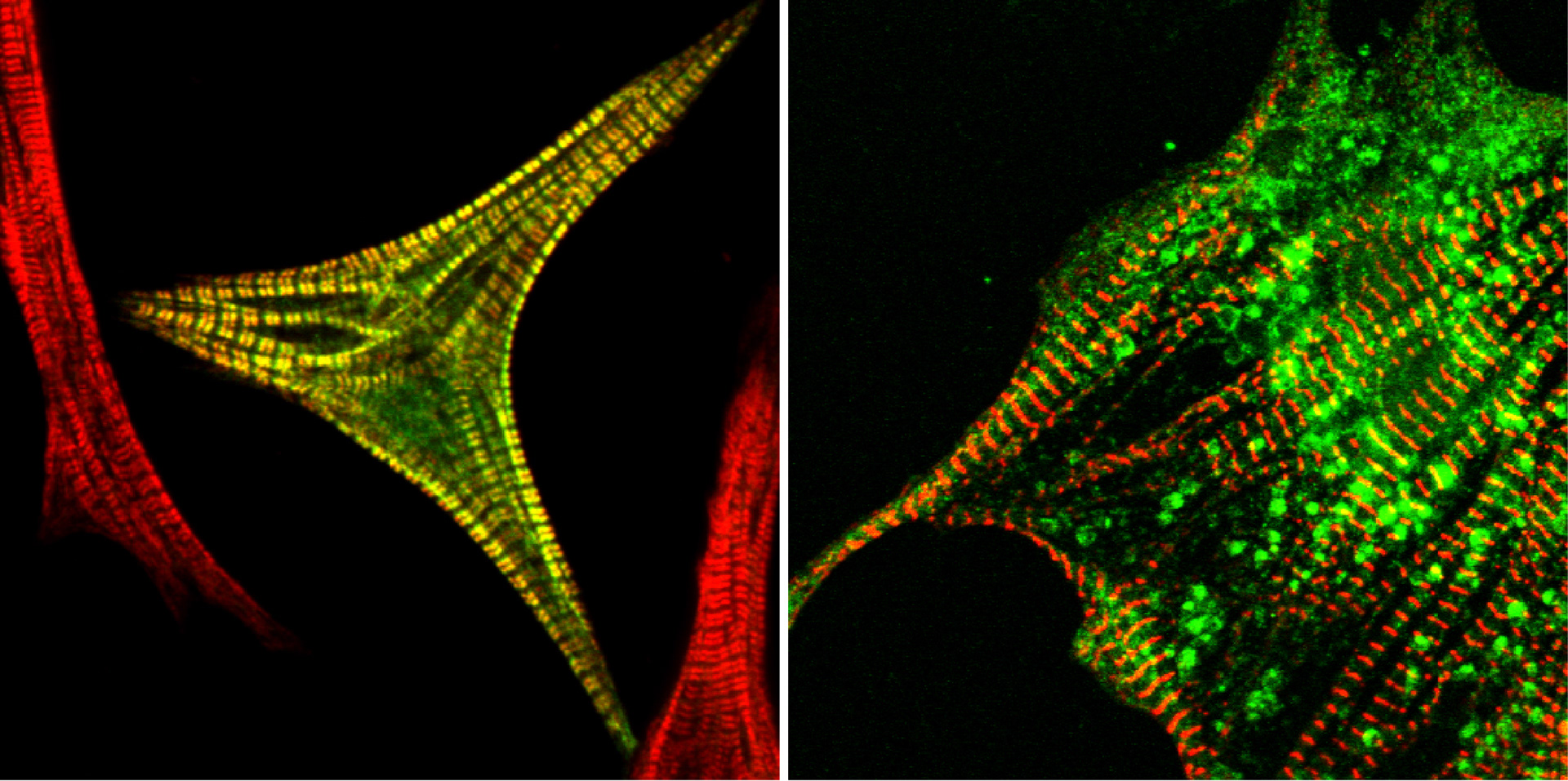
Actin Assembly Proteins in the Heart
Myofibrillogenesis is a crucial part of heart development. It results in a highly ordered array of identical multiprotein subunits, the sarcomeres. They contain actin and myosin filaments, which interact with each other to produce force.
Various models have been proposed to explain the formation of myofibrils. Depending on the model organism or cell type, studies suggested the existence of either stress fibre like structures or pre-myofibrils as precursors and backbone for myofibrillogenesis.
Regardless of the model, the initial myofibrillar structures are formed at integrin adhesion sites and contain actin filaments. However, up until now it is unclear which actin assembly proteins are involved in their formation.
Actin assembly is a very tightly regulated process with specialized assembly factors for different actin structures. Our data suggests that formin family proteins are involved in the formation of mature myofibrils at several stages and regulated by mechanical signals. Here we want to dissect when and how these proteins are recruited, turned on, or off in order to form the mature myofibrils.
Various models have been proposed to explain the formation of myofibrils. Depending on the model organism or cell type, studies suggested the existence of either stress fibre like structures or pre-myofibrils as precursors and backbone for myofibrillogenesis.
Regardless of the model, the initial myofibrillar structures are formed at integrin adhesion sites and contain actin filaments. However, up until now it is unclear which actin assembly proteins are involved in their formation.
Actin assembly is a very tightly regulated process with specialized assembly factors for different actin structures. Our data suggests that formin family proteins are involved in the formation of mature myofibrils at several stages and regulated by mechanical signals. Here we want to dissect when and how these proteins are recruited, turned on, or off in order to form the mature myofibrils.